CPC Best Plant Conservation Practices
to Support Species Survival in the Wild
to Support Species Survival in the Wild
Conventional seed banking is a fundamental plant conservation practice within the CPC network and around the world. In 1995, CPC published Guidelines for the Management of Orthodox Seeds (Wieland 1995). These guidelines not only presented practical advice for preserving the genetic diversity of seeds of the rarest plant species in North America based upon the best science of the period, they blossomed from a partnership with the ARS-USDA National Laboratory for Genetic Resources Preservation (formerly the National Seed Storage Laboratory) in Fort Collins, Colorado. The dedicated NLGRP staff, their excellent facilities, and collaborative research with CPC conservation officers have been central to our growing understanding of how to store seed while maintaining viability. While much has stayed the same, this updated version details some of the technological advances that have emerged over the last 25 years.
In addition to using recommendations from our previously published guidelines (Wieland 1995), these updated guidelines incorporate protocols from FAO’s Genebank Standards for agriculture (FAO 2014), MSBP Seed Conservation Standards (MSB 2015), our research, as well as published research from around the world. Note that the international guidelines from FAO and MSBP are broad, encompassing advice for economically important and common species, the CPC guidelines pertain to collections of rare wild species and assuring that seeds can live long enough in storage to support a conservation mandate.
Prior to making seed collections, there are several considerations a practitioner can make depending upon the type of plant material that is available. (See “Questions to Ask before Acquiring a Conservation Collection” and “Questions to Ask to Determine the Most Efficient Way to Preserve the Plant Tissue Long-Term.”) In some cases, you may need to do a reconnaissance trip to a population before you will be able to answer the questions about whether seeds are present or not. In other cases, you may need to conduct preliminary laboratory trials to determine whether seeds are viable or not, or desiccation tolerant or not.
Seeds capable of conventional freezer storage (temperatures –18°C + 3°C) are called orthodox (see “What is an ‘Orthodox’ Seed?” below). Conventional freezer storage is relatively inexpensive and highly accessible to many institutions. In most cases, the aim is to store long-term (> 20 years). If seeds are not desiccation tolerant (also known as exceptional species, recalcitrant, or intermediate; see “What Is a ‘Recalcitrant’ Seed?” and “What Is an ‘Intermediate’ Seed?” below) or you are only able to collect shoots, rather than seeds, see CPC Best Practice Chapter “Alternatives to Conventional Seed Banking”.
Several steps are required to bring seeds from the wild into the seed laboratory for processing before placing into cold storage. We cover each of these steps in this chapter which is intended to help practitioners keep orthodox seeds alive for as long as possible so that the seeds may be used for future reintroductions to the wild. We urge practitioners to follow the NEW practices to increase the longevity of seeds in storage.

A seed’s physical and physiological states determine whether it can be stored by conventional means or not. Seed water content, relative humidity and temperature are key interacting factors contributing to longevity (see Figure 1.1). In order for a seed to be stored and survive freezing temperatures, a prerequisite is that it is capable of surviving desiccation or removal of most of the water in its cells.
Research from the 1950s and 1960s demonstrated that drying and cooling crop seeds increased survival time (i.e., longevity). This response became known as “orthodox seed behavior” and is described by Harrington’s Thumb Rules, which state: (1) For each 1% decrease in moisture content, the storage life of the seed is doubled and (2) For each 10°F (5.6 °C) decrease in storage temperature, the storage life of a seed is doubled. Harrington also coined the Hundred Rule, which indicates that the storage temperature (°F) and Relative Humidity (%) should add up to less than 100 to achieve safe storage (Harrington 1960). Later Ellis and Roberts (1980) developed viability equations that predict the proportion of seeds that are viable after storage under a variety of conditions. These have been fundamental to seed conservation predictions for orthodox seeds.
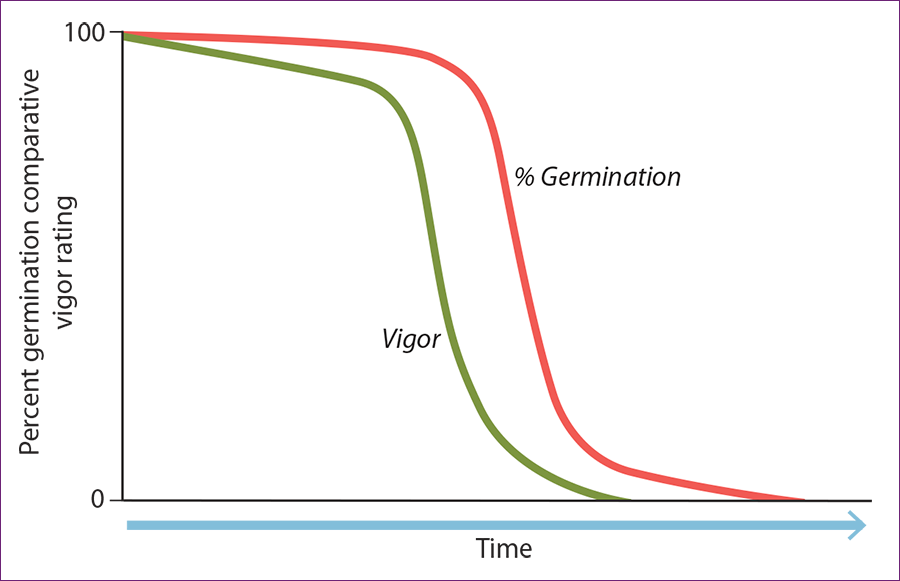
Figure 1.1 – The decline in vigor and germination of a seed accession over time. The amount of time a seed is viable in storage will be a function of seed characteristics, storage temperature and seed moisture. (Redrawn from Harrington 1960.)

Eric Roberts (1973) coined the word ‘recalcitrant’ to refer to seeds that are not orthodox. While orthodox seeds can be stored conventionally, recalcitrant seeds cannot be stored conventionally.
The term “recalcitrant” anthropomorphizes seed responses to water loss. A recalcitrant seed tolerates some water loss, but not the extreme level survived by orthodox seed. Water remaining in recalcitrant seeds forms lethal ice crystals during conventional storage.
Removing embryos from recalcitrant seeds and storing them at liquid nitrogen temperatures is a possible solution to the problem (Al-Zoubi and Normah 2015). Protocols developed for several North American species are labor-intensive but give high survival (Reed 2008). Examples of North American recalcitrant seeds that survive cryopreservation are: Zizania texana, Howellia aquatilis, Acer saccharinum, and Quercus sp. (although species native to Mexico have not yet been tested). Hawai’i and Florida may be “hotspots” within the US for seed recalcitrance, and yet we see a low incidence (<5% of species) (for example, Salazar et al. 2017).
“Recalcitrant” is also used to describe a seed that is difficult to germinate, which happens when seeds lack embryos (that is, “empty” seeds), have fastidious germination requirements (that is, dormant seeds or those with rudimentary embryos), or age quickly (possibly intermediate-type seeds). Some examples of U.S. natives that have been called recalcitrant but survive considerable desiccation are Helonias bullata, Actaea racemosa, Asimina tetramera, Castanea ozarkensis, and Magnolia ashei.

In the 1990s, it became apparent that some seeds did not fit either orthodox or recalcitrant seed categories. These seeds, known as intermediate, share functional characteristics with recalcitrant seeds and can be categorized as non-orthodox or exceptional. The viability of intermediate seeds is not maintained in the freezer and they need to be cryopreserved in liquid nitrogen. Intermediate seeds can be dried to lower water contents than recalcitrant seeds, and this means that they can be cooled at 30° to 100°C/min (compared to hundreds of degrees per sec for recalcitrant seed tissues) and still avoid lethal ice formation. Unlike recalcitrant seeds where the embryonic axis must be surgically excised to achieve required cooling rates, intermediate seeds can be placed whole into liquid nitrogen, and this process requires less labor and there is usually no impact on survival from the initial exposure to liquid nitrogen.
An early estimate of the incidence of intermediate behavior, about 10%–15% of angiosperms world-wide (Dickie and Pritchard 2002), was based on only one of several syndromes now recognized for the category (Walters 2015). Now we believe there are many more seeds that could be classified as intermediate. Moreover, the tendency to produce intermediate seeds might be a characteristic of a population and its ecology, rather than a species. Intermediate seeds were originally characterized in crop seeds—papaya (Ellis et al. 1991) and coffee (Eira et al. 2006)—but U.S. natives such as several Cuphea and Salix species also exhibit intermediate traits (Crane et al. 2006; Ballesteros and Pence 2017). We believe between 25% and 50% of endangered species from Hawai’i, especially in Campanulaceae and Gesneraceae, produce intermediate-type seeds (Walters, Weisenberger and colleagues, unpublished data).
Intermediate seeds are characterized by at least one of the following symptoms:
It is helpful to look at the categories of seed storage physiology for orthodox, recalcitrant, and intermediate seeds in the context of water content and temperature. The interaction between water content and temperature that affects the physical structure of cytoplasm is portrayed in a phase diagram, which includes a vitrification-plasticization curve (Figure 1.2). All seeds have a threshold water content, below which they are damaged by further drying (Walters 2015). At room temperature, orthodox seeds can be dried safely to water contents between 0.03 and 0.08 g H2O/g total mass (3 to 7%), while safe water content for recalcitrant seeds is above 0.20 g H2O/ g total mass (> 20%). Intermediate seeds lie in between these two levels (Figure 1.2). Below the lower thresholds, some seeds may experience increased aging and die (Walters 2015).
Examining seeds in the context of how much cell volume is occupied by water, dry matter and void space is a promising direction for research. Consider the contents of cells within a seed. As seeds mature they accumulate food reserves which replace water and reduce the water content. However the water potential (availability of water to participate in reactions) stays about the same until seeds mature and separate from the maternal plant. No longer having a supply of water, orthodox seeds dry, causing cells to shrink and molecules within the cells to compress. The proportion of cell volume occupied by dry matter at the onset of dehydration can help predict the amount of water loss that can be tolerated by a seed. See Dr. Chris Walters describe the importance of understanding the phase of water inside a cell in her 2018 keynote at the CPC National Meeting:
To preserve viability during storage, the molecules within a cell must compress until a glass forms (also called vitrification). Orthodox seeds survive to very low water contents and therefore tolerate glass formation; hence, they can be safely preserved in the freezer under conventional storage conditions. In contrast, threshold water contents for recalcitrant and intermediate seeds are higher than the water content required to form a glass. The only way to achieve a glassy state is to cool the cytoplasm profoundly. But, cooling cells containing water is dangerous because lethal ice crystals will likely form. Therefore, preservation methods must cool seeds rapidly to avoid ice formation. For recalcitrant seeds, this can be on the order of hundreds of degrees per second, and it may require excising the embryo to make the sample small enough to cool quickly enough. Once the glass forms, the material must be stored at liquid nitrogen temperatures to avoid warming and inducing ice formation. Because rare species’ seeds come in all shapes, sizes, and physiologies, we present this physical chemistry perspective to help practitioners understand challenges to preservation and make wise decisions for their seeds.
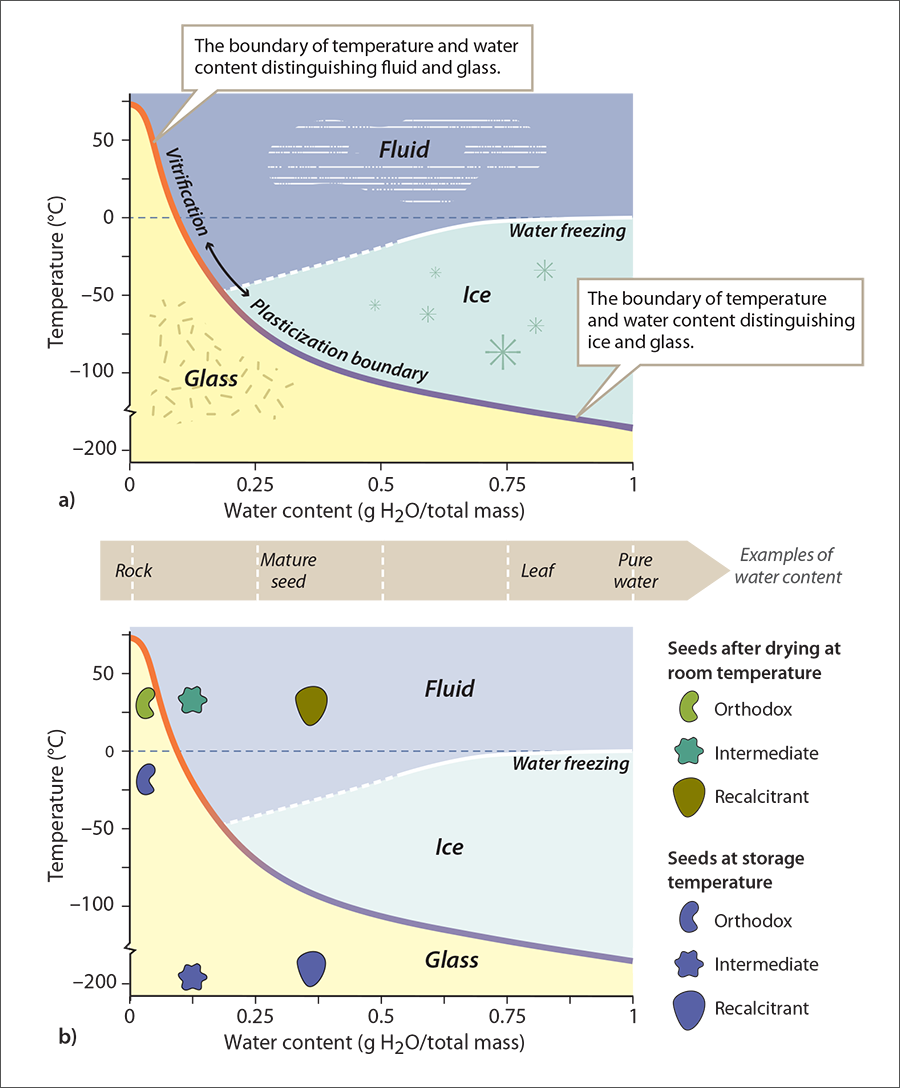
Figure 1.2 – Orthodox, recalcitrant and intermediate seeds and the vitrification-plasticization curve. a) Phase diagram showing boundaries between fluid, ice and glass in relation to temperature and water content. b) Seeds categorized as orthodox, intermediate, or recalcitrant are dried to different water contents. To preserve viability, seeds must be dried and cooled. Note that at conventional freezer temperatures (–20°C), orthodox seeds are in glass, however intermediate and recalcitrant seeds must be cooled rapidly to liquid nitrogen temperatures to be in glass and avoid ice formation.
Baskin, C. M., and J. M. Baskin. 2014. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2nd ed. Academic Press, San Diego.
Brown, A. D. H., and D. R. Marshall. 1995. A basic sampling strategy: theory and practice. Collecting plant genetic diversity: technical guidelines. CABI, Wallingford, UK: 75–91.
Bureau of Land Management. 2016. Technical protocol for the collection, study, and conservation of seeds from native plant species for Seeds of Success.
Falk, D. A., and K. E. Holsinger. 1991. Genetics and conservation of rare plants. New York, Oxford University Press.
Food and Agriculture Organization of the United Nations (FAO). 2014. Genebank standards for plant genetic resources for food and agriculture. Rome, Italy. http://www.fao.org/3/a-i3704e.pdf.
Gordon, D., and C. Gantz. 2008. Screening new plant introductions for potential invasiveness: a test of impacts for the United States. Conservation Letters 1:227–235.
Gordon, D. R., D. A. Onderdonk, A. M. Fox, and R. K. Stocker. 2008a. Consistent accuracy of the Australian weed risk assessment system across varied geographies. Diversity and Distribution 14:234–242.
Gordon, D. R, D. A. Onderdonk, A. M. Fox, R. K. Stocker, and C. Gantz. 2008b. Predicting invasive plants in Florida using the Australian weed risk assessment. Invasive Plant Science and Management 1:176–195.
Griffth, M.P., M. Calonje, A. W. Meerow, F. Tut, A.T. Kramer, A. Hird, T.M. Magellan, and C.E. Husby. 2015. Can a botanic garden cycad collection capture the genetic diversity in awild population. Int. J. Plant Science 176: 1-10.
Guerrant, E. O., Jr., and P. L. Fiedler. 2004. Accounting for sample decline during ex situ storage and reintroduction. Pages 365–385 in E. O. Guerrant, Jr., K. Havens, and M. Maunder, editors. Ex situ plant conservation: supporting species survival in the wild. Island Press, Washington, DC.
Guerrant, E. O., Jr., K. Havens, and M. Maunder, editors. 2004. Ex situ plant conservation: supporting species survival in the wild. Island Press, Washington, DC.
Millennium Seed Bank Partnership (MSB). 2015. Seed conservation standards for “MSB Partnership Collections.” Royal Botanic Gardens, Kew, UK.
Reichard, S., H. Liu, and C. Husby. 2012. Is managed relocation of rare plants another pathway for biological invasions? In J. Maschinski and K. E. Haskins, editors. Plant reintroduction in a changing climate: promises and perils. Island Press, Washington, DC.
Seed Conservation Hub. Accessed August 3, 2017..
Volk, G. M., D. R. Lockwood, and C. M. Richards. 2007. Wild plant sampling strategies: the roles of ecology and evolution. In Plant breeding reviews, volume 29. John Wiley and Sons, Inc., New York, New York.
Wieland, G. D. 1995. Guidelines for the management of orthodox seeds. Center for Plant Conservation, St. Louis.
Wyse, S. V., and J. B. Dickie. 2016. Predicting the global incidence of seed desiccation sensitivity. Journal of Ecology. doi: 10.1111/1365-2745.12725.
 Daniel Ballesteros
Daniel Ballesteros
 Kim Taylor
Kim Taylor
 Christina Walters
Christina Walters
 Cheryl Birker
Cheryl Birker
 Joyce Maschinski
Joyce Maschinski
 Christina Walters
Christina Walters 


